Part I Biology: Introduction
Lithops, when a person first sees them, are ugly plants. But it's bizarre appearance and more bizarre desire to seemingly never want water, surviving in the wild for 20+ months between rain, is endearing. Lithops can complete an entire deciduous cycle of it's single two succulent leaves dying, and two new leaves growing to adult size without any external water. (Water recycling in leaves of Lithops (Aizoaceae). Otto et al. 2013). This unique adaptation, in addition to a contractile tap root which draws the drying plant into the soil, lends the plant to its common name "Living stones".
It was first seen by Europeans in 1811: "An picking up from the ground, what was supposedly a curiously shaped pebble, it proved to be a plant, and an additional new species to the numerous tribe of the Mesembryanthemem but in colour and appearance bore the closest resemblance to the stones between which it was growing" (Burchell 1821).
I was first introduced to a package of seeds of Lithops spp. while working at a greenhouse in secondary school. I was told of the difficulties people seemed to have germinating them, and though the idea of "living stones" was interesting, the thought of failing put me off these plants for several years.
Interest was sparked again this year via joining the succulent enthusiasts go-to-forum "reddit.com/r/succulents". As well, I was fortunate enough to acquire the excellent, and well researched volume by a prolific Lithops enthusiast and macro photographer Harald Jainta, and his extremely skilled partner Anja Jainta, which suggested the grassland species L. aucampiae and L. lesliei can tolerate living with grasses, which made me hopeful to experiment with growing them.

The first Lithops you may see might be labelled "Lithops", if you're lucky. Species are very rarely marked at garden centres, or hardware stores, and even experts seem stumped and have to resort to cutting the adult plants in half to speciate. Lithops are considered by most to be challenging to speciate. They have very limited morphological characteristics (seeds, capsules), and the flowers, a common morphological tool for taxonomists are nearly identical between species.
In 1922 the Lithops were placed in a larger group Mesembryanthemums (cf. Conophytum, Dinteranthus, Titanopsis). Even now these plants are affectionately referred to as mesembs. The initial speciation of the genus "Lithops" differentiated the genus based on flower colour (Xantholithops vs. Leucolithops viz. Schwantes 1957), and the presence of light conducting windows on the flat tops of the succulent leaves (Fenestratae vs. Afenestratae viz. Nel. 1946). Window pattern analysis ballooned the number of species to over 90. However Cole (1979) considered the windowing and leaf colour to be more of a selection pressure by herbivores, and Lithops to be limited to no more than 40 species.
Today it's believed that different species can appear nearly identical in leaf colour. Lithops that mimic endemic rocks/soil are more robust to predation. Recall the classic biology story of the absence of white moths in soot stained London, while black varieties had a survival benefit, and were well and good in London. A professor of African Languages, Desmond Cole, and partner Naureen Cole collected extensively and published many papers on Lithops, and attempted to resolve the taxonomy of Lithops. Robert Wallace (1988) did his Ph.D. dissertation on a systemic review of Lithops that avoided subjective pattern recognition on Lithops leaves.
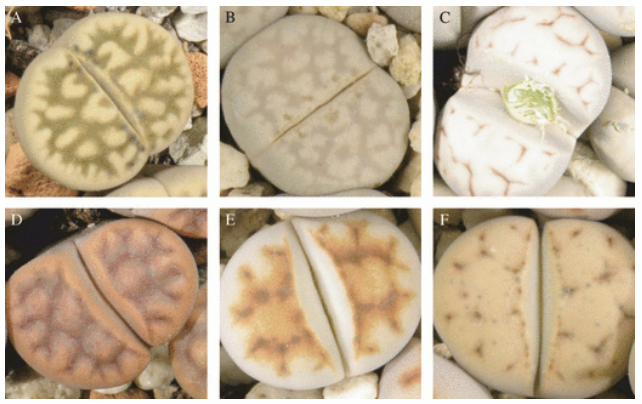
Lithops karasmontana varieties (Kellner et al. 2011)
Lithops Biology:
Summary of Wallace 1988 Dissertation in regards to taxonomy

Lithops cross-section (Wallace, 1988)
TI: Colourations which can occur in various places, CL: Cleft, CP: thin walled water storage tissues with large central vacuole, PV: Primary vein, CHL: photosynthetic tissues, P: apical and axilliary meristem, R: Root
Lithops are composed of two fleshy succulent leaves which are fused or bifurcated, at the base. The stem cells, the meristem, is located above a single tap-root. "Windows" on the upper leaf surface, have thinner waxy cuticle than the rest of the plant, and the cell outer walls are thinner. These epidermal cells are convex at the air/plant interface which may help concentrate light deeper in the plant.
Populations found today are very cryptic, in that they are nearly identical to surrounding rocks. They are eaten by Springbok antelope, and grazing sheep or goats. Luckily, the position of the meristem at the base of the plant provides them the opportunity to grow new sets of leaves and even flower after their adult leaves are eaten.
In general the body is underground with only the flat leaf tops visible. They grow in gravelly or rocky medium derived from granite, sandstone, limestone, quartzite, gneiss, pegmatite, or schist, etc. Plants that are covered by soil also lose less moisture to transpiration, and are known to be frost resistant in dry or porous soils.
Oxygen and carbon dioxide are exchanged through pairs of stomata guard cells which are on all skin tissue, but most densely concentrated near the base of the plant. Only one species is unique in regards to stomata in that it has three stomata guard cells, Lithops dorotheae. If the top of the leaves were entirely green, light would not penetrate very deeply. Lithops has adapted to dessication pressure by limiting the exposure to air, and developing its photosynthetic tissue in the underground portions adjacent to it's transparent windows. Nel described the plant as a mine shaft with light entering the mouth of the mine and lighting the interior (1946).
The main xylem and phloem tissues often curve from the base, around the central region into the top most central cleft, and downward. This may represent an evolutionary origin of the current morphology. Cross-sections of the plant suggest stem facing epidermal tissues are greatly reduced or adsorbed to facilitate light penetration to chloroplasts deep in the plant. In the evolution of Lithops, the proximal tissues could have fused, folded, and the surface then truncated to the current flat leaf surface.
The main contractile root is permanent and will pull a drying plant into the ground while secondary and tertiary roots die-back then rapidly regrow when moisture is present. This is similar to a convergent evolutionary strategy in cacti, which also have deciduous secondary roots with high salt concentrations to collect water by osmosis. Experiments in the early 20th century show that the entire plant, including old roots are nearly impervious to water. A plant weighing 23.95 g absobed 3.3 g water in 15 days. This same plant absorbed 7 g of water when a small cut was made at the base of the plant and immersed in water.

Lithops flower cross-section (Wallace, 1988)
S: Style, ST: Stamen, PT: Petal, N: Nectary, SE: Septal, I: Colouration and calcium oxalate raphides, R: Recepticle, PE: Pedicel, FS: False septum, P: Placenta, PS: Placental septum, TS: True septum.
Pollen is most commonly longer on the polar axis, however in some species the pollen is near spherical, or even longer on the equatorial axis. All pollen is microverrucate, or slightly textured. Apertures are oblong, and in some species no perforations of the subsurface exine layer are seen, and more commonly microperforations are present in the subsurface of the pollen viewed through apertures.
Seed capsules retain seed until moistened, and shaped in such a way that raindrops will carry seeds down and away from the parent plant. Studies have shown seeds will drop within 20 cm of the parent 75% of the time, but have been observed to carry up to 75 cm from the parent plant. Seeds do not float, but populations have been seen in linear formations down gentle slope suggesting water dispersal during heavy rain events. As the Lithops grow in specialised environments long-distance dispersal would be less likely to result in germination than local dispersal where a population already exists. Three species are known to only one locality (i.e. L. werneri, L. viridis [200 m<sup>2</sup> only], L. helmutii) and critically endangered. As for the very wide geographic distribution of several species (i.e. L. lesliei, L. verruculosa) are not well explained, and speculated to be carried on avian feet, or on the hairs of herbavores muzzles or feet.

Lithops seed cross-section (Wallace, 1988)
RP: Rostral papillum, Ro: Rostrum, Ra: Radicle, Em: Embryo, CP: Cotyledon pair (first leaves), FR: Funicular remnant (umbilical), SC: Seed coat, En: Endosperm (energy store).

Lithops lesliei moistened seed © Atreyu 2018.
Seed coat has torn mid-seed/endosperm as seed swells with moisture. Slight swelling noticeable to the rostrum.
Seeds range from 0.35 mm (i.e. L. verruculosa, L. marmorata) to 1.50 mm (Lithops lesliei), and germinate rapidly, with best results anecdotally reported after at least one year from production. The tearing of the seed coat is not used for taxonomy, however it is reported that some species carry the seed coat on the edge of the cotyledon (i.e. L. lesliei, L. pseudotruncalis).
Seedlings grow as a fused cotyledons, with a cleft in the centre of the fusion, or across the entire surface of the plant. Primary leaves most commonly take 6 - 12 months to emerge from the cotyledon, though in my own experience Lithops karasmontana primary leaves started to emerge after 12 - 14 weeks from germination.

Lithops karasmontana primary leaves emerging. © Atreyu 2018
Note: cotyledon appears slightly pink/red compared to emerging primary leaves.
After flowering is initiated at the apical meristem, leaves will cycle annually from growth initiated at the axilliary meristem, with one pair of leaves. Two pairs of leaves can grow if each axilliary meristem initiate. Some species will have only one "head" form with one axilliary meristem remaining inactive each year (i.e. monocephalic species, cf. Lithops gracilidelineata). As the old leaves age, water is transferred to the developing leaves, and has been documented in fluoroscopic studies. Old dead leaves form a protective film on the body of the new leaves, and act as a heat shield from incident sunlight, and to protect from abrasion to the rough soils.
Lithops Genetics
As summarized in a genetic analysis (Kellner et al. 2011), the Lithops subfamily evolved over a 3-8 million year period. Adaptations included cellular features such as wider water conducting cells in the xylem without perforations, and seed capsules that would only open when moist.
Similar to most mesebryanthemums, Lithops are diploid with 9 chromosomes (2n=18), with only one exception in Lithops dinteri v brevis (2n=17). Additional genetic analysis is needed, as current analysis have been limited to enzyme electrophoresis, or just one or two plant or chloroplast genes.
Lithops show high inter-fertility between all the yellow flowered species, and separately between all the white flowered species. However, white flowered variants of yellow flowered species are common, suggesting the biochemical pathway for colouration is controlled by few genes. This may also be the case with plant colouration due the very broad plant colouration, even in single species.
Coming soon Part II: Growth Experiments 4 Months Deep.
No comments:
Post a Comment